Know your neuron: Grid cells March 3, 2008
Posted by Johan in Animals, Know your neuron, Navigation, Neuroscience.3 comments
Know your neuron is a mini-series here at the Fan Club where we look at some of the curious response characteristics of neurons discovered through electrophysiology. We will work our way through the simple properties of cells that respond to spatial locations or head direction and on to increasingly abstract cells, which encode the identity of individuals or perhaps even the relation between actions of others and those of the self.
![]() Given how straight-forward their response properties are, it is curious that grid cells were only discovered relatively recently (Hafting et al, 2005). According to one of my professors, this is simply because researchers didn’t put their rats in sufficiently large boxes to see the pattern previously, but who knows… The response properties of grid cells are perhaps best illustrated in a figure (this is from the excellent Scholarpedia article on Grid cells, written by two of their discoverers).
Given how straight-forward their response properties are, it is curious that grid cells were only discovered relatively recently (Hafting et al, 2005). According to one of my professors, this is simply because researchers didn’t put their rats in sufficiently large boxes to see the pattern previously, but who knows… The response properties of grid cells are perhaps best illustrated in a figure (this is from the excellent Scholarpedia article on Grid cells, written by two of their discoverers).
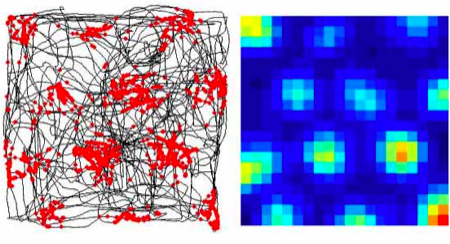
In this figure, the x and y coordinates correspond to the spatial location of a rat, which is running around freely inside a large box. The black lines in the left figure shows how this particular rat explored the box in a fairly haphazard manner. However, an electrode inserted in the rat’s subcortex picks up a signal that is anything but chaotic: the responses of said neuron are given as red dots in the left figure, while the right figure gives the firing rate distribution (ranging from blue for silent and red for the peak rate of responding). Although the rat is running about randomly, this neuron is responding in a grid, seemingly coming on an off in response to the animal’s spatial location.
This response pattern is similar regardless of the exact box, so the pattern does not reflect learning of the spatial layout of this box as such. Indeed, when the light is turned out, the grid cell continues to respond as before while the rat runs around in the dark (Hafting et al, 2005 – this is not terribly impressive when you consider how little rats use vision, however). Yet, at least some grid cells showed a response pattern that rotated 90 degrees with a corresponding shift in an external visual cue, so although the grid cells do not depend on the external environment, they do seem to take cues from it when available.
Grid cells are found in Entorhinal cortex of the rat, a region which provides the main input to the Hippocampus, long known as a centre for memory consolidation and spatial navigation. Thus, these cells are a prime candidate as building blocks for more complex spatial cells that are observed in the Hippocampus proper. Different grid cells in Entorhinal cortex appear to have partially overlapping response fields, so that a given location might produce a response in some grid cells but not others. There is also a topographical structure, where the size of the grid increases as one moves from the dorsomedial to the ventrolateral side of the area. Incidentally, this pattern matches a spatial scale increase of more complex cells in the Hippocampus proper.
The obvious implication is that by combining the output of a number of partially-overlapping grid cells, perhaps a cell with more complex spatial properties could be built. The next issue of Know your neuron is about one category of such neurons, which appear to respond only to a specific spatial location.
References
Hafting, T., Fyhn, M., Molden, S., Moser, M., & Moser, E.I. (2005). Microstructure of a spatial map in the entorhinal cortex. Nature, 436, 801-806. DOI: 10.1038/nature03721
Moser, E., & Moser, M.-B. (2007). Grid Cells. Scholarpedia, 2, 3394.
Learning to recognise faces: perceptual narrowing? January 11, 2008
Posted by Johan in Animals, Developmental Psychology, Face Perception, Sensation and Perception.add a comment

![]() That image certainly piques your interest, doesn’t it? Sugita (2008) was interested in addressing one of the ancient debates in face perception: the role of early experience versus innate mechanisms. In a nutshell, some investigators hold that face perception is a hardwired process, others that every apparently special face perception result can be explained by invoking the massive expertise we all possess with faces, compared to other stimuli. Finally, there is some support for a critical period during infancy, where a lack of face exposure produces irreparable face recognition deficits (see for example Le Grand et al, 2004). Unfortunately, save for a few unfortunate children who are born with cataracts, there is no real way to address this question in humans.
That image certainly piques your interest, doesn’t it? Sugita (2008) was interested in addressing one of the ancient debates in face perception: the role of early experience versus innate mechanisms. In a nutshell, some investigators hold that face perception is a hardwired process, others that every apparently special face perception result can be explained by invoking the massive expertise we all possess with faces, compared to other stimuli. Finally, there is some support for a critical period during infancy, where a lack of face exposure produces irreparable face recognition deficits (see for example Le Grand et al, 2004). Unfortunately, save for a few unfortunate children who are born with cataracts, there is no real way to address this question in humans.
Enter the monkeys, and the masked man. Sugita (2008) isolated monkeys soon after birth, and raised them in a face-free environment for 6, 12 or 24 months. After this, the monkeys were exposed to strictly monkey or human faces for an additional month.
At various points during this time, Sugita (2008) tested the monkeys on two tasks that were originally pioneered in developmental psychology as means of studying pre-lingual infants. In the preferential looking paradigm, two items are presented, and the time spent looking at either item in the pair is recorded. The monkeys viewed human faces, monkey faces, and objects, in various combinations. It is assumed that the monkey (or infant) prefers whichever item it looks at more. In the paired-comparison procedure, the monkey is primed with the presentation of a face, after which it views a face pair, where one of the faces is the same as that viewed before. If the monkey views the novel face more, it is inferred that the monkey has recognised the other face as familiar. So the preferential looking paradigm measures preference between categories, while the paired-comparison procedure measures the ability to discriminate items within a category.
Immediately following deprivation, the monkeys showed equal preference for human and monkey faces. By contrast, a group of control monkeys who had not been deprived of face exposure showed a preference for monkey faces. This finding suggests that at the very least, the orthodox hard-wired face perception account is wrong, since the monkeys should then prefer monkey faces even without previous exposure to them.
In the paired-comparison procedure, the control monkeys could discriminate between monkey faces but not human faces. By contrast, the face-deprived monkeys could discriminate between both human and monkey faces. This suggests the possibility of perceptual narrowing (the Wikipedia article on it that I just linked is probably the worst I’ve read – if you know this stuff, please fix it!), that is, a tendency for infants to lose their ability to discriminate between categories which are not distinguished in their environment. The classic example occurs in speech sounds, where infants can initially discriminate phoneme boundaries (e.g., the difference between /bah/ and /pah/ in English) that aren’t used in their own language, although this ability is lost relatively early on in the absence of exposure to those boundaries (Aslin et al, 1981). But if this is what happens, surely the face-deprived monkeys should lose their ability to discriminate non-exposed faces, after exposure to faces of the other species?
Indeed, this is what Sugita (2008) found. When monkeys were tested after one month of exposure to either monkey or human faces, they now preferred the face type that they had been exposed to over the other face type and non-face objects. Likewise, they could now only discriminate between faces from the category they had been exposed to.
Sugita (2008) didn’t stop there. The monkeys were now placed in a general monkey population for a year, where they had plenty of exposure to both monkey and human faces. Even after a year of this, the results were essentially identical as immediately following the month of face experience. This implies that once the monkeys had been tuned to one face type, that developmental door was shut, and no re-tuning occurred. Note that in this case, one month of exposure to one type trumped one year of exposure to both types, which shows that as far as face recognition goes, what comes first seems to matter more than what you get the most of.
Note a little quirk in Sugita’s (2008) results – although the monkeys were face-deprived for durations ranging from 6 to 24 months, these groups did not differ significantly on any measures. In other words, however the perceptual narrowing system works for faces, it seems to be flexible about when it kicks in – it’s not a strictly maturational process that kicks in at a genetically-specified time. This conflicts quite harshly with the cataract studies I discussed above, where human infants seem to lose face processing ability quite permanently when they miss out on face exposure in their first year. One can’t help but wonder if Sugita’s (2008) results could be replicated with cars, houses, or any other object category instead of faces, although this is veering into the old ‘are faces special’ debate… It’s possible that the perceptual narrowing observed here is a general object recognition process, unlike the (supposedly) special mechanism with which human infants learn to recognise faces particularly well.
On the applied side, Sugita (2008) suggests that his study indicates a mechanism for how the other-race effect occurs – that is, the advantage that most people display in recognising people of their own ethnicity. If you’ve only viewed faces of one ethnicity during infancy (e.g., your family), perhaps this effect has less to do with racism or living in a segregated society, and more to do with perceptual narrowing.
References
Sugita, Y. (2008). Face perception in monkeys reared with no exposure to faces. Proceedings of the National Academy of Sciences (USA), 105, 394-398.
